Кто является представителем утопического социализма. Экономическое учения социалистов-утопистов. Промышленный класс будущего
Читайте также
Развивался таким образом, что функция каждой его системы стала результатом функции суммы клеток, из которых состоят органы и ткани данной системы. Каждая клетка организма располагает набором структур и механизмов, позволяющих ей осуществлять собственный метаболизм и выполнять присущую ей функцию.
В состав клетки входят цитоплазматическая или поверхностная мембрана; цитоплазма, имеющая ряд органелл, включений, элементов цитоскелета; ядро, содержащее ядерный геном. Органеллы клетки и ядро отграничены в цитоплазме внутренними мембранами. Каждая структура клетки выполняет в ней свою функцию, а все они вместе взятые обеспечивают жизнеспособность клетки и выполнение ею специфических функций.
Ключевая роль в осуществлении клеточных функций и их регуляции принадлежит цитоплазматической мембране клетки.
Общие принципы строения цитоплазматической мембраны
Для всех клеточных мембран характерен один принцип строения (рис. 1), в основе которого лежат физико-химические свойства сложных липидов и белков, входящих в их состав. Мембраны клетки располагаются в водной среде и для понимания физико-химических явлений, влияющих на их структурную организацию, полезным является описание взаимодействия липидных и белковых молекул с молекулами воды и друг с другом. Ряд свойств клеточных мембран также вытекает из рассмотрения этого взаимодействия.
Известно, что плазматическая мембрана клетки представлена двойным слоем сложных липидов, покрывающим поверхность клетки на всем ее протяжении. Для создания липидного бислоя в ее структуру могли быть отобраны природой и включены только те молекулы липидов, которые обладают амфифильными (амфипатическими) свойствами. Этим условиям отвечают молекулы фосфолипидов и холестерола. Их свойства таковы, что одна часть молекулы (глицерольная для фосфолипидов и циклопентановая для холестерола) обладает полярными (гидрофильными) свойствами, а другая (жирнокислотные радикалы) — неполярными (гидрофобными) свойствами.
Рис. 1. Строение цитоплазматической мембраны клетки.
Если определенное количество молекул фосфолипидов и холестерола поместить в водную среду, то они спонтанно начнут собираться в упорядоченные структуры и формировать замкнутые пузырьки (липосомы ), в которых оказывается заключенной часть водной среды, а поверхность становится покрытой непрерывным двойным слоем (бислоем ) фосфолипидных молекул и холестерола. При рассмотрении характера пространственного расположения молекул фосфолипидов и холестерола в этом бислое видно, что молекулы данных веществ располагаются своими гидрофильными частями в сторону наружного и внутреннего водных пространств, а гидрофобными — в противоположных направлениях — внутрь бислоя.
Что заставляет молекулы этих липидов самопроизвольно формировать в водной среде бислойные структуры, подобные структуре бислоя клеточной мембраны? Пространственное расположение амфифильных молекул липидов в водной среде диктуется одним из требований термодинамики. Наиболее вероятной пространственной структурой, которую сформируют в водной среде молекулы липидов, будет структура, обладающая минимумом свободной энергии .
Такой минимум свободной энергии в пространственной структуре липидов в воде будет достигнут в случае, когда и гидрофильные, и гидрофобные свойства молекул будут реализованы в виде соответствующих межмолекулярных связей.
При рассмотрении поведения сложных амфифильных молекул липидов в воде можно объяснить и некоторые свойства клеточных мембран . Известно, что если механически повредить плазматическую мембрану (например, проколоть ее электродом или через прокол удалить ядро и поместить в клетку другое ядро), то через мгновение за счет сил межмолекулярного взаимодействия липидов и воды мембрана самопроизвольно восстановит целостность . Под действием таких же сил можно наблюдать слияние бислоев двух мембран при их соприкосновении (например, везикул и пресинаптической мембраны в синапсах). Способность мембран сливаться при их непосредственном контакте является частью механизмов обновления структуры мембран, транспорта компонентов мембран из одного субклеточного пространства в другое, а также частью механизмов эндо- и экзоцитоза.
Энергия межмолекулярных связей в липидном бислое очень низкая, поэтому создаются условия для быстрого перемещения в мембране молекул липидов и белков и для изменения структуры мембраны при воздействии на нее механических сил, давлений, температуры и других факторов. Наличие в мембране двойного липидного слоя образует замкнутое пространство, изолирует цитоплазму от окружающей водной среды и создаст препятствие для свободного прохождения воды и растворимых в ней веществ через клеточную мембрану. Толщина липидного бислоя составляет около 5 нм.
В состав клеточных мембран также входят белки. Их молекулы по объему и массе в 40-50 раз больше, чем молекулы мембранных липидов. За счет белков толщина мембраны достигает 7-10 нм. Несмотря на то что суммарные массы белков и липидов в большинстве мембран почти равны, количество молекул белков в мембране в десятки раз меньше, чем молекул липидов.
Что же произойдет, если белковая молекула окажется помещенной в фосфолипидный бислой липосом, наружные и внутренние поверхности которых полярны, а внутрилипидный неполярен? Под влиянием сил межмолекулярных взаимодействий липидов, белка и воды произойдет формирование такой пространственной структуры, в которой неполярные участки пептидной цепи будут стремиться расположиться в глубине липидного бислоя, в то время как полярные — занять положение на одной из поверхностей бислоя и могут к тому же оказаться погруженными во внешнюю или внутреннюю водную среду липосомы. Очень сходный характер расположения белковых молекул имеет место и в липидном бислое клеточных мембран (рис. 1).
Обычно белковые молекулы локализуются в мембране разрозненно одна от другой. Возникающие в неполярной части бислоя липидов очень слабые силы гидрофобных взаимодействий между углеводородными радикалами липидных молекул и неполярными участками белковой молекулы (липид-липидные, липид-белковые взаимодействия) не препятствуют протеканию процессов тепловой диффузии этих молекул в структуре бислоя.
Когда с помощью тонких методов исследования была изучена структура клеточных мембран, то оказалось, что она очень сходна с той, которая самопроизвольно формируется фосфолипидами, холестеролом и белками в водной среде. В 1972 г. Синджером и Никольсом была предложена жидкостно-мозаичная модель строения клеточной мембраны и сформулированы ее основные принципы.
Согласно этой модели, структурную основу всех клеточных мембран составляет жидкоподобный непрерывный двойной слой амфипатических молекул фосфолипидов, холестсрола, гликолипидов, самопроизвольно формирующих его в водной среде. В липидном бислое асимметрично расположены белковые молекулы, выполняющие специфические рецепторные, ферментативные и транспортные функции. Белковые и липидные молекулы обладают подвижностью и могут совершать вращательные движения, диффундировать в плоскости бислоя. Белковые молекулы способны изменять их пространственную структуру (конформацию), смещаться и изменять свое положение в липидном бислое мембраны, погружаясь на различную глубину или всплывая на его поверхность. Структура липидного бислоя мембраны неоднородна. В нем имеются участки (домены), получившие название «рафты», которые обогащены сфинголипидами и холестеролом. «Рафты» отличаются фазовым состоянием от состояния остальной части мембраны, в которой они располагаются. Особенности строения мембран зависят от выполняемой ими функции и функционального состояния.
Исследование состава клеточных мембран подтвердили, что основными их компонентами являются липиды, составляющие около 50% массы плазматической мембраны. Около 40-48% массы мембраны приходится на белки и 2-10% — на углеводы. Остатки углеводов либо входят в состав белков, образуя гликопротеины, либо липидов, образуя гликолипиды. Фосфолипиды являются главными структурными липидами плазматических мембран и составляют 30-50% их массы.
Углеводные остатки молекул гликолипидов обычно располагаются на внешней поверхности мембраны и погружены в водную среду. Они играют важную роль в межклеточных, клеточно-матриксных взаимодействиях и распознавании антигенов клетками иммунной системы. Молекулы холестерола, встроенные в фосфолипидный бислой, способствуют сохранению упорядоченного расположения жирнокислотных цепей фосфолипидов и их жидкокристаллического состояния. В связи с наличием высокой конформационной подвижности ацильных радикалов жирных кислот фосфолипидов они формируют достаточно рыхлую упаковку липидного бислоя и в нем могут образовываться структурные дефекты.
Белковые молекулы способны пронизывать всю мембрану так, что их концевые участки выступают за се поперечные пределы. Такие белки называют трансмембранными , или интегральными . В составе мембран имеются также белки, только частично погруженные в мембрану или располагающиеся на ее поверхности.
Многие специфические функции мембран определяются белковыми молекулами, для которых липидная матрица является непосредственным микроокружением и от ее свойств зависит осуществление функций белковыми молекулами. Среди важнейших функций мембранных белков можно выделить: рецепторную — связывание с такими сигнальными молекулами, как нейромедиаторы, гормоны, ингерлейкины, факторы роста, и передача сигнала на пострецепторные структуры клетки; ферментативную — катализ внутриклеточных реакций; структурную — участие в формировании структуры самой мембраны; транспортную — перенос веществ через мембраны; каналообразующую — формирование ионных и водных каналов. Белки совместно с углеводами участвуют в осуществлении адгезии-слипания, склеивания клеток при иммунных реакциях, объединении клеток в слои и ткани, обеспечивают взаимодействие клеток с внеклеточным матриксом.
Функциональная активность мембранных белков (рецепторов, ферментов, переносчиков) определяется их способностью легко изменять свою пространственную структуру (конформацию) при взаимодействии с сигнальными молекулами, действии физических факторов или изменении свойств среды микроокружения. Энергия, требующаяся для осуществления этих конформационных изменений структуры белков, зависит как от внутримолекулярных сил взаимодействия отдельных участков пептидной цепи, так и от степени текучести (микровязкости) мембранных липидов, непосредственно окружающих белок.
Углеводы в виде гликолипидов и гликопротеинов составляют лишь 2-10% от массы мембраны; количество их в разных клетках изменчиво. Благодаря им осуществляются некоторые виды межклеточных взаимодействий, они принимают участие в узнавании клеткой чужеродных антигенов и совместно с белками создают своеобразную антигенную структуру поверхностной мембраны собственной клетки. По таким антигенам клетки узнают друг друга, объединяются в ткань и на короткое время слипаются для передачи сигнальных молекул друг другу.
Благодаря низкой энергии взаимодействия входящих в мембрану веществ и относительной упорядоченности их расположения клеточная мембрана приобретает ряд свойств и функций, не сводимых к простой сумме свойств образующих ее веществ. Незначительные по силе воздействия на мембрану, сравнимые с энергией межмолекулярных связей белков и липидов, могут вести к изменению конформации белковых молекул, проницаемости ионных каналов, изменению свойств мембранных рецепторов, других многочисленных функций мембраны и самой клетки. Высокая чувствительность структурных компонентов плазматической мембраны имеет решающее значение в восприятии клеткой информационных сигналов и их преобразовании в ответные реакции клетки.
Функции цитоплазматической мембраны клетки
Цитоплазматическая мембрана выполняет многие функции, обеспечивающие жизненные потребности клетки и, в частности, ряд функций необходимых для восприятия и передачи клеткой информационных сигналов.
Среди важнейших функций плазматической мембраны можно выделить:
- отграничение клетки от окружающей се среды с сохранением формы, объема и существенных различий между клеточным содержимым и внеклеточным пространством;
- перенос веществ внутрь клетки и из нее на основе свойства избирательной проницаемости, активного и других видов транспорта;
- поддержание трансмембранной разности электрических потенциалов (поляризации мембраны) в покое, ее изменение при различных воздействиях на клетку, генерация и проведение возбуждения;
- участие в обнаружении (рецепции) сигналов физической природы, сигнальных молекул за счет формирования сенсорных или молекулярных рецепторов и передаче сигналов внутрь клетки;
- образование межклеточных контактов (плотный, щелевой и десмосомальный контакт) в составе образуемых тканей или при адгезии клеток различных тканей;
- создание гидрофобного микроокружения для проявления активности ферментов, связанных с мембраной;
- обеспечение иммунной специфичности клетки за счет наличия в структуре мембраны антигенов белковой или гликопротеиновой природы. Иммунная специфичность имеет значение при объединении клеток в ткань и взаимодействии с клетками, осуществляющими иммунный надзор в организме.
Приведенный перечень функций клеточных мембран свидетельствует о том, что они принимают участие в осуществлении не только клеточных функций, но и базисных процессов жизнедеятельности органов, тканей и целостного организма. Без знания ряда явлений и процессов, обеспечиваемых мембранными структурами, невозможно понимание и осознанное выполнение некоторых диагностических процедур и лечебных мероприятий. Например, для правильного применения многих лекарственных веществ необходимо знание того, в какой мере каждое из них проникает через клеточные мембраны из крови в тканевую жидкость и в клетки.
Цитоплазматическая мембрана (плазмалемма) — основная, универсальная для всех клеток часть поверхностного аппарата. Ее толщина составляет около 10 нм. Плазмалемма ограничивает цитоплазму и защищает ее от внешних воздействий, принимает участие в процессах обмена веществ между клеткой и внеклеточной средой.
Основными компонентами мембраны являются липиды и белки. Липиды составляют около 40 % массы мембран. Среди них преобладают фосфолипиды.
Молекулы фосфолипидов располагаются в виде двойного слоя (липидный бислой). Как вы уже знаете, каждая молекула фосфолипида образована полярной гидрофильной головкой и неполярными гидрофобными хвостами. В цитоплазматической мембране гидрофильные головки обращены к наружной и внутренней сторонам мембраны, а гидрофобные хвосты — внутрь мембраны (рис. 30).
Кроме липидов, в состав мембран входят белки двух типов: интегральные и периферические. Интегральные белки более или менее глубоко погружены в мембрану либо пронизывают ее насквозь. Периферические белки располагаются на внешней и внутренней поверхностях мембраны, причем многие из них обеспечивают взаимодействие плазмалеммы с надмембранными и внутриклеточными структурами.
На внешней поверхности цитоплазматической мембраны могут располагаться молекулы олиго- и полисахаридов. Они ковалентно связываются с мембранными липидами и белками, образуя гликолипиды и гликопротеины. В клетках животных такой углеводный слой покрывает всю поверхность плазмалеммы, образуя надмембранный комплекс. Он называется гликокаликсом (от лат. гликис — сладкий, калюм — толстая кожа).
Функции цитоплазматической мембраны. Плазмалемма выполняет ряд функций, важнейшими из которых являются барьерная, рецепторная и транспортная.
Барьерная функция. Цитоплазматическая мембрана окружает клетку со всех сторон, играя роль барьера — преграды между сложно организованным внутриклеточным содержимым и внеклеточной средой. Барьерную функцию обеспечивает, прежде всего, липидный бислой, не позволяющий содержимому клетки растекаться и препятствующий проникновению в клетку чужеродных веществ.
Рецепторная функция. В цитоплазматическую мембрану встроены белки, способные в ответ на действие различных факторов внешней среды изменять свою пространственную структуру и таким образом передавать сигналы внутрь клетки. Следовательно, цитоплазматическая мембрана обеспечивает раздражимость клеток (способность воспринимать раздражители и определенным образом реагировать на них), осуществляя обмен информацией между клеткой и окружающей средой.
Некоторые рецепторные белки цитоплазматической мембраны способны распознавать определенные вещества и специфически связываться с ними. Такие белки могут участвовать в отборе необходимых молекул, поступающих в клетки.
К рецепторным белкам относятся, например, антигенраспознающие рецепторы лимфоцитов, рецепторы гормонов и нейромедиаторов и т. д. В осуществлении рецепторной функции, кроме мембранных белков, важную роль играют элементы гликокаликса.
Разнообразие и специфичность наборов рецепторов на поверхности клеток приводит к созданию сложной системы маркеров, позволяющих отличать s.свои:/ клетки (той же особи или того же вида) от s.чужих:/. Благодаря этому клетки могут вступать друг с другом во взаимодействия (например, конъюгация у бактерий, образование тканей у животных).
В цитоплазматической мембране могут быть локализованы специфические рецепторы, реагирующие на различные физические факторы. Например, в плазмалемме светочувствительных клеток животных расположена специальная фоторецепторная система, ключевую роль в функционировании которой играет зрительный пигмент родопсин. С помощью фоторецепторов световой сигнал превращается в химический, что, в свою очередь, приводит к возникновению нервного импульса.
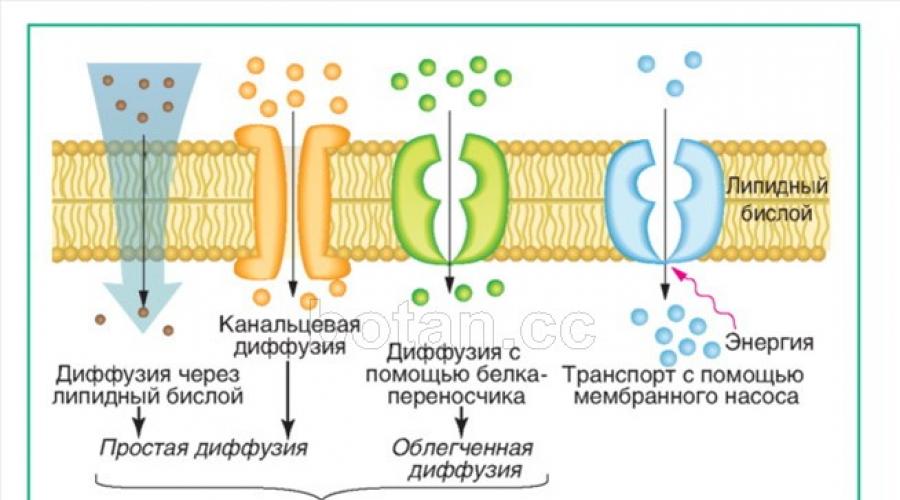
Транспортная функция. Одной из основных функций плазмалеммы является обеспечение транспорта веществ как в клетку, так и из нее во внеклеточную среду. Выделяют несколько основных способов транспорта веществ через цитоплазматическую мембрану: простая диффузия, облегченная диффузия, активный транспорт и транспорт в мембранной упаковке (рис. 31).
При простой диффузии наблюдается самопроизвольное перемещение веществ через мембрану из области, где концентрация этих веществ выше, в область, где их концентрация ниже. Путем простой диффузии через плазмалем-му могут проходить небольшие молекулы (например, Н 2 0, 0 2 , С0 2 , мочевина) и ионы. Как правило, неполярные вещества транспортируются непосредственно через липидный бислой, а полярные молекулы и ионы — через каналы, образованные специальными мембранными белками. Простая диффузия происходит относительно медленно. Для ускорения диффузного транспорта существуют мембранные белки-переносчики. Они избирательно связываются с тем или иным ионом или молекулой и переносят их через мембрану. Такой тип транспорта называется облегченной диффузией. Скорость переноса веществ при облегченной диффузии во много раз выше, чем при простой.
Диффузия (простая и облегченная) — разновидности пассивного транспорта. Он характеризуется тем, что вещества транспортируются через мембрану без затрат энергии и только в том направлении, где наблюдается меньшая концентрация данных веществ.

Активный транспорт — перенос веществ через мембрану из области низкой концентрации этих веществ в область более высокой. Для этого в мембране имеются специальные насосы, работающие с использованием энергии (см. рис. 31). Чаще всего для работы мембранных насосов используется энергия АТФ.
Одним из наиболее распространенных мембранных насосов является натрий-калиевая АТ Фаза (Na + /K + - АТ Фаза). Она удаляет из клетки ионы Na + и закачивает в нее ионы К + - Для работы Ыа + /К + -АТФаза использует энергию, выделяемую при гидролизе АТФ. Благодаря этому насосу поддерживается разность концентраций Na + и К + в клетке и внеклеточной среде, что лежит в основе многих биоэлектрических и транспортных процессов.
В результате активного транспорта с помощью мембранных насосов происходит также регуляция содержания Mgr + , Са 2+ и других ионов в клетке.
Путем активного транспорта через цитоплазматическую мембрану могут перемещаться не только ионы, но и моносахариды, аминокислоты, другие низкомолекулярные вещества.
Своеобразной и относительно хорошо изученной разновидностью мембранного транспорта является транспорт в мембранной упаковке. В зависимости от того, в каком направлении переносятся вещества (в клетку или из нее), различают два вида этого транспорта — эндоцитоз и экзоцитоз.
Эндоцитоз (отгреч. эндон — внутри, китос — клетка, ячейка) — поглощение клеткой внешних частиц путем образования мембранных пузырьков. При эндоцитозе определенный участок плазмалеммы обволакивает внеклеточный материал и захватывает его, заключая в мембранную упаковку (рис. 32).
Выделяют такие разновидности эндоцитоза, как фагоцитоз (захват и поглощение твердых частиц) и пиноцитоз (поглощение жидкости).
Путем эндоцитоза осуществляется питание гетеротрофных протистов, защитные реакции организма (поглощение лейкоцитами чужеродных частиц) и др.
Экзоцитоз (от греч. экзо — снаружи) — транспортировка веществ, заключенных в мембранную упаковку, из клетки во внешнюю среду. Например, пузырек комплекса Гольджи перемещается к цитоплазматической мембране и сливается с ней, а содержимое пузырька выделяется во внеклеточную среду. Таким способом клетки выделяют пищеварительные ферменты, гормоны и другие вещества.
1. Можно ли увидеть плазмалемму в световой микроскоп? Каковы химический состав " и строение цитоплазматической мембраны?
2. Что такое гликокаликс? Для каких клеток он характерен?
3. Перечислите и поясните основные функции плазмалеммы.
4. Какими способами может осуществляться транспорт веществ через мембрану? В чем заключается принципиальное отличие пассивного транспорта от активного?
5. Чем отличаются процессы фагоцитоза и пиноцитоза? В чем проявляется сходство этих процессов?
6. Сравните различные типы транспорта веществ в клетку. Укажите черты их сходства и различия.
7. Какие функции не смогла бы выполнять цитоплазматическая мембрана, если бы в ее состав не входили белки? Ответ обоснуйте.
8. Некоторые вещества (например, диэтиловый эфир, хлороформ) проникают через биологические мембраны даже быстрее, чем вода, хотя их молекулы намного больше молекул воды. С чем это связано?
- § 1. Содержание химических элементов в организме. Макро- и микроэлементы
- § 2. Химические соединения в живых организмах. Неорганические вещества
- § 10. История открытия клетки. Создание клеточной теории
- § 15. Эндоплазматическая сеть. Комплекс Гольджи. Лизосомы
- § 24. Общая характеристика обмена веществ и преобразование энергии
Глава 1. Химические компоненты живых организмов
Глава 2. Клетка - структурная и функциональная единица живых организмов
Глава 3. Обмен веществ и преобразование энергии в организме
Глава 4. Структурная организация и регуляция функций в живых организмах
Цитоплазма — обязательная часть клетки, заключенная между плазматической мембраной и ядром; подразделяется на гиалоплазму (основное вещество цитоплазмы), органоиды (постоянные компоненты цитоплазмы) и включения (временные компоненты цитоплазмы). Химический состав цитоплазмы: основу составляет вода (60-90% всей массы цитоплазмы), различные органические и неорганические соединения. Цитоплазма имеет щелочную реакцию. Характерная особенность цитоплазмы эукариотической клетки — постоянное движение (циклоз ). Оно обнаруживается, прежде всего, по перемещению органоидов клетки, например хлоропластов. Если движение цитоплазмы прекращается, клетка погибает, так как, только находясь в постоянном движении, она может выполнять свои функции.
Гиалоплазма (цитозоль ) представляет собой бесцветный, слизистый, густой и прозрачный коллоидный раствор. Именно в ней протекают все процессы обмена веществ, она обеспечивает взаимосвязь ядра и всех органоидов. В зависимости от преобладания в гиалоплазме жидкой части или крупных молекул, различают две формы гиалоплазмы: золь — более жидкая гиалоплазма и гель — более густая гиалоплазма. Между ними возможны взаимопереходы: гель превращается в золь и наоборот.
Функции цитоплазмы:
- объединение всех компонентов клетки в единую систему,
- среда для прохождения многих биохимических и физиологических процессов,
- среда для существования и функционирования органоидов.
Клеточные оболочки
Клеточные оболочки ограничивают эукариотические клетки. В каждой клеточной оболочке можно выделить как минимум два слоя. Внутренний слой прилегает к цитоплазме и представлен плазматической мембраной (синонимы — плазмалемма, клеточная мембрана, цитоплазматическая мембрана), над которой формируется наружный слой. В животной клетке он тонкий и называется гликокаликсом (образован гликопротеинами, гликолипидами, липопротеинами), в растительной клетке — толстый, называется клеточной стенкой (образован целлюлозой).
Все биологические мембраны имеют общие структурные особенности и свойства. В настоящее время общепринята жидкостно-мозаичная модель строения мембраны . Основу мембраны составляет липидный бислой, образованный в основном фосфолипидами. Фосфолипиды — триглицериды, у которых один остаток жирной кислоты замещен на остаток фосфорной кислоты; участок молекулы, в котором находится остаток фосфорной кислоты, называют гидрофильной головкой, участки, в которых находятся остатки жирных кислот — гидрофобными хвостами. В мембране фосфолипиды располагаются строго упорядоченно: гидрофобные хвосты молекул обращены друг к другу, а гидрофильные головки — наружу, к воде.
Помимо липидов в состав мембраны входят белки (в среднем ≈ 60%). Они определяют большинство специфических функций мембраны (транспорт определенных молекул, катализ реакций, получение и преобразование сигналов из окружающей среды и др.). Различают: 1) периферические белки (расположены на наружной или внутренней поверхности липидного бислоя), 2) полуинтегральные белки (погружены в липидный бислой на различную глубину), 3) интегральные, или трансмембранные, белки (пронизывают мембрану насквозь, контактируя при этом и с наружной, и с внутренней средой клетки). Интегральные белки в ряде случаев называют каналообразующими, или канальными, так как их можно рассматривать как гидрофильные каналы, по которым в клетку проходят полярные молекулы (липидный компонент мембраны их бы не пропустил).

А — гидрофильная головка фосфолипида; В — гидрофобные хвостики фосфолипида; 1 — гидрофобные участки белков Е и F; 2 — гидрофильные участки белка F; 3 — разветвленная олигосахаридная цепь, присоединенная к липиду в молекуле гликолипида (гликолипиды встречаются реже, чем гликопротеины); 4 — разветвленная олигосахаридная цепь, присоединенная к белку в молекуле гликопротеина; 5 — гидрофильный канал (функционирует как пора, через которую могут проходить ионы и некоторые полярные молекулы).
В состав мембраны могут входить углеводы (до 10%). Углеводный компонент мембран представлен олигосахаридными или полисахаридными цепями, связанными с молекулами белков (гликопротеины) или липидов (гликолипиды). В основном углеводы располагаются на наружной поверхности мембраны. Углеводы обеспечивают рецепторные функции мембраны. В животных клетках гликопротеины образуют надмембранный комплекс — гликокаликс, имеющий толщину несколько десятков нанометров. В нем располагаются многие рецепторы клетки, с его помощью происходит адгезия клеток.
Молекулы белков, углеводов и липидов подвижны, способны перемещаться в плоскости мембраны. Толщина плазматической мембраны — примерно 7,5 нм.
Функции мембран
Мембраны выполняют такие функции:
- отделение клеточного содержимого от внешней среды,
- регуляция обмена веществ между клеткой и средой,
- деление клетки на компартаменты («отсеки»),
- место локализации «ферментативных конвейеров»,
- обеспечение связи между клетками в тканях многоклеточных организмов (адгезия),
- распознавание сигналов.
Важнейшее свойство мембран — избирательная проницаемость, т.е. мембраны хорошо проницаемы для одних веществ или молекул и плохо проницаемы (или совсем непроницаемы) для других. Это свойство лежит в основе регуляторной функции мембран, обеспечивающей обмен веществ между клеткой и внешней средой. Процесс прохождения веществ через клеточную мембрану называют транспортом веществ . Различают: 1) пассивный транспорт — процесс прохождения веществ, идущий без затрат энергии; 2) активный транспорт — процесс прохождения веществ, идущий с затратами энергии.
При пассивном транспорте вещества перемещаются из области с более высокой концентрацией в область с более низкой, т.е. по градиенту концентрации. В любом растворе имеются молекулы растворителя и растворенного вещества. Процесс перемещения молекул растворенного вещества называют диффузией, перемещения молекул растворителя — осмосом. Если молекула заряжена, то на ее транспорт влияет и электрический градиент. Поэтому часто говорят об электрохимическом градиенте, объединяя оба градиента вместе. Скорость транспорта зависит от величины градиента.
Можно выделить следующие виды пассивного транспорта: 1) простая диффузия — транспорт веществ непосредственно через липидный бислой (кислород, углекислый газ); 2) диффузия через мембранные каналы — транспорт через каналообразующие белки (Na + , K + , Ca 2+ , Cl -); 3) облегченная диффузия — транспорт веществ с помощью специальных транспортных белков, каждый из которых отвечает за перемещение определенных молекул или групп родственных молекул (глюкоза, аминокислоты, нуклеотиды); 4) осмос — транспорт молекул воды (во всех биологических системах растворителем является именно вода).
Необходимость активного транспорта возникает тогда, когда нужно обеспечить перенос через мембрану молекул против электрохимического градиента. Этот транспорт осуществляется особыми белками-переносчиками, деятельность которых требует затрат энергии. Источником энергии служат молекулы АТФ. К активному транспорту относят: 1) Na + /К + -насос (натрий-калиевый насос), 2) эндоцитоз, 3) экзоцитоз.

Работа Na + /К + -насоса . Для нормального функционирования клетка должна поддерживать определенное соотношение ионов К + и Na + в цитоплазме и во внешней среде. Концентрация К + внутри клетки должна быть значительно выше, чем за ее пределами, а Na + — наоборот. Следует отметить, что Na + и К + могут свободно диффундировать через мембранные поры. Na + /К + -насос противодействует выравниванию концентраций этих ионов и активно перекачивает Na + из клетки, а K + в клетку. Na + /К + -насос представляет собой трансмембранный белок, способный к конформационным изменениям, вследствие чего он может присоединять как K + , так и Na + . Цикл работы Na + /К + -насоса можно разделить на следующие фазы: 1) присоединение Na + с внутренней стороны мембраны, 2) фосфорилирование белка-насоса, 3) высвобождение Na + во внеклеточном пространстве, 4) присоединение K + с внешней стороны мембраны, 5) дефосфорилирование белка-насоса, 6) высвобождение K + во внутриклеточном пространстве. На работу натрий-калиевого насоса тратится почти треть всей энергии, необходимой для жизнедеятельности клетки. За один цикл работы насос выкачивает из клетки 3Na + и закачивает 2К + .
Эндоцитоз — процесс поглощения клеткой крупных частиц и макромолекул. Различают два типа эндоцитоза: 1) фагоцитоз — захват и поглощение крупных частиц (клеток, частей клеток, макромолекул) и 2) пиноцитоз — захват и поглощение жидкого материала (раствор, коллоидный раствор, суспензия). Явление фагоцитоза открыто И.И. Мечниковым в 1882 г. При эндоцитозе плазматическая мембрана образует впячивание, края ее сливаются, и происходит отшнуровывание в цитоплазму структур, отграниченных от цитоплазмы одиночной мембраной. К фагоцитозу способны многие простейшие, некоторые лейкоциты. Пиноцитоз наблюдается в эпителиальных клетках кишечника, в эндотелии кровеносных капилляров.
Экзоцитоз — процесс, обратный эндоцитозу: выведение различных веществ из клетки. При экзоцитозе мембрана пузырька сливается с наружной цитоплазматической мембраной, содержимое везикулы выводится за пределы клетки, а ее мембрана включается в состав наружной цитоплазматической мембраны. Таким способом из клеток желез внутренней секреции выводятся гормоны, у простейших — непереваренные остатки пищи.
Перейти к лекции №5 «Клеточная теория. Типы клеточной организации»
Перейти к лекции №7 «Эукариотическая клетка: строение и функции органоидов»